Overview of Biophysical Interactions
Biophysical Interactions Involving Phytoplankton
Biophysical Interactions Involving Zooplankton
Biophysical Interactions Involving Benthic Invertebrates
Biophysical Interactions Involving Marine Fish and Shellfish
Biophysical Interactions Involving Bird Populations
Summary of Biophysical Interactions-Hypothesis Formulation
In the CCS the underlying importance of large scale physical forcing on marine organisms is evidenced in several comparative studies. Hollowed et al. (1987) noted that extreme year classes of several marine fish tend to occur synchronously even when species are separated by large geographical distances. Synchronicity over large geographic areas would not be expected if the principal processes influencing survival were linked to local ocean conditions. These synchronous extreme year classes tend to occur during periods of unusual environmental conditions (Hollowed and Wooster 1992, Beamish 1993). Further evidence of the importance of large scale physical forcing on marine organisms is found in numerous single species studies that demonstrate an apparent relationship between survival and physical forcing (Tables 7-8). Many of these studies show apparent linkages between biological response variables and temperature, wind driven transport or ENSO. Long-term trends in the abundance of marine fish stocks also show coherent patterns of association with large scale physical forcing. Francis and Sibley (1991) and Beamish and Boullion (1993) found coherent long term population trends in many stocks of Pacific salmon, with the southern (CCS) and northern (Canadian and Alaskan) stocks being out of phase. On an even broader scale, time series of Pacific sardine catch from populations in the southeast, northeast, and northwest Pacific all show remarkably similar trends (Kawasaki 1992). These comparative studies suggest that production of marine fish stocks is linked to large scale (but unknown) ocean processes.
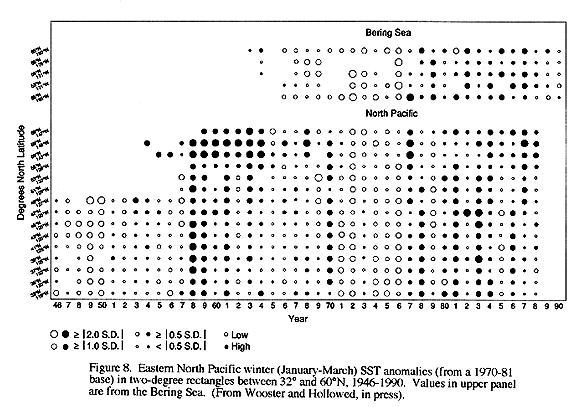
A notable feature of time series of physical and biological measurements in the eastern boundary currents are that they both exhibit abrupt changes in state. The most recent shift occurred in the mid 1970s when coastal sea surface temperatures abruptly became warmer than average in the California Current (Hollowed and Wooster 1992, Trenberth 1990, Royer 1993, Figure 8). This shift was accompanied by a drop in zooplankton abundance, recovery of the depleted Pacific sardine population in the Southern California, and reduced size of many salmon runs in Oregon and Washington. Evidence of similar shifts at earlier times are found in the paleoclimatic records (Baumgartner et al. 1992). Together the paleoclimate and historic records suggest that the CCS response to gradually increasing concentrations of greenhouse gasses may be abrupt changes in qualitative states of the physical and biological system, rather than gradual change.
A principal goal of the U.S. GLOBEC CCS project is to identify the processes linking physical change to observed biological responses at the population level. Quantifying the impact of environmental change on survival rates of marine organisms may be difficult because the responses are often non-linear and may involve a sequence of events rather than a single factor. Detailed studies of the processes underlying survival have been conducted for only a few fish/shellfish species (English sole, northern anchovy, salmon, Dungeness crab, sea urchins, shortbelly and widow rockfish (underway)), the common barnacle, the copepod Calanus marshallae, and a euphausiid (Euphausia pacifica) in the CCS. These studies have illustrated the importance of environmental processes that occur on the scale of days, weeks and seasons. Sources of short term variability on the scale of days to weeks include upwelling-relaxation events, changes in turbulent mixing of the upper ocean and variability in upwelling jets and California Current meanders and eddies. Variations on seasonal scales include changes in storm tracks and frequency, frequency and intensity of upwelling, turbulent mixing, poleward flow (Davidson Current), California Current position and strength, air-sea heat exchange and freshwater runoff.
This overview illustrates that while there appears to be coherence in time series of abundance on the long time and large space scale, there is also considerable variability on the short time and small space scales. Predicting the consequences of climate change ultimately rests on accounting for the processes that generate variation in population processes at all time and space scales.
The following section summarizes our current understanding of the major biophysical interactions in each region. This background may assist in the development of research initiatives designed to enhance or understanding of processes influencing production of our nations marine resources in the CCS.
There appears to be a close link between mesoscale features (eddies, jets) and the distribution of marine organisms in Region II. Satellite imagery suggests high concentrations of chlorophyll are associated with jets, eddies and other mesoscale features that occur as a result of coastal upwelling and the advection of this water (Flament et al. 1985). Chavez et al. (1991) found that high-nutrient and phytoplankton regions occur south and inshore of the Pt. Arena upwelling jet. Two distinct phytoplankton communities (coastal diatom and oceanic single-cell) are found in Region II and occur as separate populations on either side of upwelling fronts or divided by jets (Chavez et al. 1991).
In Region III, dense dinoflagellate blooms, often with very restricted vertical and horizontal distributions, are common features of the Southern California Bight during the summer (reviewed in Beers, 1986). Many of these dinoflagellates are important prey for larval anchovies (e.g. Lasker et al., 1970). Lasker (1981) noted differences in the abundance and nutritional content of dinoflagellates and diatoms in Region III as related to the food requirements of first feeding northern anchovy. Lasker's work demonstrated the importance of identifying not only the amount of production, but the type of production that occurs in a given year.
Within a season, several studies indicate climate change could influence zooplankton by influencing the location and persistence of mesoscale features such as semipermanent eddies, or upwelling jets. In Region I, consistent maintenance of some copepod species within a nearshore band has been explained on the basis of two-celled upwelling ( Peterson, et al. 1979) and intermittent upwelling (Wroblewski 1980).
Zooplankton in Region II appear to be influenced by mesoscale features. Smith and Lane (1991) hypothesize that upwelling jets or eddies may transport the copepod, Eucalanus californicus, to the surface and onshore where egg laying occurs. Mackas et al. (1991) summarized the distribution of zooplankton near a jet in Region II. This study showed zooplankton species composition can change dramatically in response to fronts, resulting in distinct coastal and oceanic zooplankton communities. More recently, Laurs, Lynn and Peterson (unpublished) conducted a CTD, chlorophyll and zooplankton survey of a portion of the California Current jet and two associated eddies (34 deg -37 deg N, 123 deg -126 deg W, several hundred kilometers off Monterey and Point Sur) in July 1991. The Mackas survey and the Laurs et al. survey demonstrated that the California Current was enriched in zooplankton hundreds of kilometers seaward of the coast, in contrast to Washington and Oregon where zooplankton biomass is far higher on the shelf than over the slope. The zone of enrichment is separated from the warm offshore water by the meandering California Current "jet", a narrow (50 km wide) zone of low salinity water (32.7-32.9 ppt) that winds its way southward from the Oregon shelf to as far as southern California.
Numerous studies have been conducted on biophysical interactions of zooplankton in Region III. The distributions of many zooplankton taxa in Region III are influenced by a recurrent, deep offshore eddy southwest of Point Conception. The distributions of euphausiids, both those species with cool water, more northerly affinities and those with warm, more southerly affinities have been shown to be markedly affected by this offshore eddy (Haury et al. 1986), as have several other zooplankton species (Haury 1984).
Spatial variations in primary production and microplankton food influence different taxa in different ways. For example, in Region III, Checkley (1980) found that the copepod Paracalanus parvus became increasingly more food limited with distance from shore (to 50 km), while Calanus pacificus and Rhincalanus nasutus were found to have higher lipid contents and presumably better nutritional conditions in offshore waters of the CCS (Hakanson 1984). On smaller spatial scales, multi-species patches of copepods occur (Star and Mullin 1979), suggesting some common patterns of response to patch-generating processes.
Earlier (Section IV) we developed hypothetical climate change scenarios based on our current understanding of the processes linking climate and oceanography. Sixteen key physical changes were identified (Table 1). In this section we extend our hypotheses to include potential direct impacts of these changes to major fish and shellfish communities. This exercise is not intended to suggest that we are certain of the biological responses. The importance of the exercise is to identify the most important biophysical interactions and to point out areas where we lack knowledge about specific biological responses to physical changes.
Based on the background information found in Tables 7-8 and the predicted influence of climate change (Table 1) we identified the expected biophysical responses of marine fish and shellfish (Tables 9-11[Region I fish and shellfish; Region III fish]). An important outcome of this exercise was that it revealed areas where the expected biological responses were unknown. Very few studies have been conducted to identify the impact of changes in turbulent mixing (variable 9), transport in eddies and meanders (variable 10), changes in the depth of the thermocline (variable 13), or changes in the intensity or frequency of storms (variables 14 and 15).
Many species exhibited mixed responses to the climate change scenario. The most common case of mixed responses was the expected biological response to upwelling and temperature. This mixed response may be a product of our hypothesized physical scenario, where both upwelling-favorable winds (based on the Bakun hypothesis) and temperatures increase (based on general warming in GCMs). Increased upwelling may still produce cooler temperatures at the coast than farther offshore, but the mean temperature will be everywhere warmer than at present. If the hypothesized increase in stratification is strong enough or if the thermocline is deeper, the upwelled water may come from above the thermocline and nutricline and decrease both the cooling and fertilizing effect of upwelling, as in the warm phase of ENSO events off South America. Since most of the biophysical studies were conducted under present ocean conditions, an inverse correlation between upwelling and temperature is built into our present thinking. The presence of mixed reponses to upwelling and surface temperatures illustrates the need for an understanding of the process underlying the biological responses, particularly when the relationship between physical variables is expected to change. Use of comparative studies between latitudinal regions, between several EBCs, and measurements during unusual periods (such as warm ENSO events) will help broaden our experience and understanding.
The predicted response of marine fish and shellfish to our hypothetical climate change scenario is consistent with the ideas of Bailey and Incze (1985) who expected that species at the northern (southern) end of their range would increase (decrease) in response to El Nino like ocean conditions. The majority of the species that spawn in Region III showed positive responses to our global warming scenario (Table 11). The notable exception to this trend is the northern anchovy stock that is expected to decline (Table 11). Predicting the response of species that spawn in Region I is more difficult because of the large number of unknown responses (Table 9-10). However, the tables suggest that coho and chinook salmon, Pacific herring and dungeness crab stocks will all decline.
The importance of developing an understanding of processes underlying production of marine fish and shellfish has been recognized and process oriented research has been initiated for a few key species. In Region I, process oriented research has focused on larval transport mechanisms. In Region I, direct observations of spatial distribution of some planktonic organisms, and the temporal course of settlement give some indication of potential transport mechanisms. In observations of the cross-shore distribution off the Oregon coast, pelagic larvae (primarily fish) could be separated into 3 zones: coastal (<30 km), transitional (approximately 30 km), and offshore (> 30 km) (Richardson and Pearcy 1977, Richardson, et al. 1980). These were thought to be due to the distance from shore of spawning locations and the fact that currents were primarily alongshore. Some crabs and fish which migrate into the neuston were exceptions, being spawned nearshore but appearing offshore. During the 1982-83 ENSO, most of the ichthyoplankton found nearshore were "offshore" species (Brodeur et al. 1985). Hobbs et al. (1992) showed that the cross shelf distribution of Dungeness crab megalopae over the California to Washington coast matched that expected on the basis of a wind driven transport mechanism.
In Region II strong offshore flows (jets) may lead to short nearshore residence times. Species that rely on advection to return to nearshore locales may not survive in this environment. Eggs and larvae will be carried offshore in regions where the coastal jet first separates from the coast (flowing from shelf to deep ocean) or meanders back onto or near the shelf and then offshore again. At present, species that successfully spawn in Region II brood their eggs and larvae or extrude live larvae (e.g., rockfishes), or use protected embayments as spawning and nursery grounds (e.g., Pacific herring) to reduce the probability of offshore transport of pelagic larval stages (Parrish et al. 1981). Wing et al. (in press) examined event scale processes and found that crab larvae (primarily Cancer sp.) settled reliably in response to upwelling relaxation. Concentration of larvae in upwelling shadows south of promontories, then subsequent northward transport may provide a retention mechanism in the face of strong southward currents during upwelling. Offshore transport also may affect larvae by the removal of their prey.
Region III is the main spawning ground for the major pelagic species of the CCS (Pacific sardine, northern anchovy, Pacific hake, Jack mackerel and Pacific mackerel). While process oriented studies have been conducted on many species in Region III, the most complete study has been conducted on the northern anchovy. NOAA's SWFC conducted process oriented research on the northern anchovy for the last twenty five years. These studies include analysis of the spatial distribution of spawning (MacCall 1984), temperature effects on adult maturation (Fiedler et al. 1986), estimates of the rates of egg production (Stauffer and Picquelle 1981), larval retention through transport or eddies (Power 1986, Hewitt 1981, Fiedler 1986), egg dispersal and incubation rates (Smith and Hewitt 1985, Zwiefel and Lasker 1981), larval condition (O'Connell 1980, Theilacker 1986, Theilacker and Watanabe 1989), larval daily growth and mortality rates (Methot 1983, Butler 1989, Lo 1986), predator distributions (Hunter and Kimbrell 1980, Webb 1981, Alvarino 1980), the influence of turbulence on prey type and availability (Scura and Jerde 1977, Lasker 1975, 1978, Wroblewski 1984) and regional differences in survival (Owen et al. 1989). These studies demonstrate that factors determining year class strength of pelagic spawning marine fish are complicated and can involve several different physical events (or processes).
The most extensive research linking oceangraphic processes to production and survival of marine birds has been conducted in Region II. In this region a unique 22-year time series of data has been collected on the reproductive phenology, demography, diet and at-sea occurrence patterns of several species including the Cassin's Auklet, a planktivorous seabird. This time series shows the auklet to be a sensitive indicator of the effects of coastal upwelling on zooplankton abundance and species composition, especially the euphausiids, Thysanoessa spinifera (a coastal species) and Euphausia pacifica (an oceanic species). The auklets near the Farallon Islands breed successfully in years when T. spinifera is available in great quantity near the surface and poorly or not at all when the euphausiid is not available, as during ENSO warming events (Tables 12 and 13; Ainley and Boekelheide 1990). Egg laying closely follows the "spring transition" to upwelling conditions (Huyer et al. 1979), which often leads rapidly to the offshore spread of cold, upwelled water and surface swarms of T. spinifera. During late summer, when T. spinifera is no longer available as prey (as demonstrated by its disappearance and the appearance of Euphausia pacifica in the auklet diet), breeding ceases and remaining chicks are abandoned.
The importance of the auklet lies both in their use as zooplankton samplers (through diet analysis) and as analogs for other top trophic level species. They are especially useful because their favored prey, T. spinifera, is also important in the diet of other species including targets of major fisheries (hake, rockfish and salmon), other sea birds and baleen whales. For instance, analysis of stomach contents of salmon show that they, like the auklet, switch in spring from their winter diet of cephalopods and other prey to gorge on T. spinifera in the outer waters of Region II (Adams et al., ms).
{kind=link}