Predictions from Historical Data
Predictions from Models
Predictions from First Principles
Climate Change Scenarios
Several long time series have been used to document the natural physical and biological variability in the CCS. These include shore-based and merchant vessel measurements (~100 years); subsurface biological and physical measurements from the CalCOFI data set (45 years) and additional surveys from specific regional projects (various dates); fish otolith collections (documenting annual growth over the past 60-100 years); and high resolution varved sediments from the Santa Barbara Basin (fish scales, diatoms, foraminifera and radiolaria over the past 1500 years). These time series show distinct quasi-periodic variability in the physical and biological components of the ecosystem, with interannual to interdecadal time scales. Moreover, ocean warming has already been measured in the California Current using hydrographic time-series data from the CalCOFI program. Roemmich (1992) documented a general ocean warming of ca. 0.8 deg C and a rise in sea level between 1950 and 1990 along the southwest U.S. coast. The data indicate that the warming was greatest in the upper 100 m, but was significant to depths of 300 m.
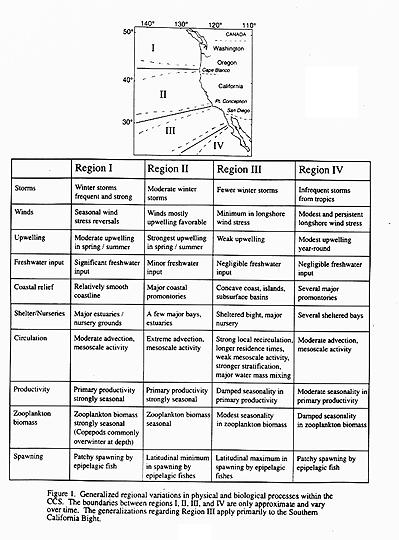
The largest source of variability at the interannual (2-10 year) time scale is the ENSO cycle. Following the warm phase in the tropics, the observed effects at mid-latitude reach the coastal ocean by both an oceanic and an atmospheric path, causing warming of the upper ocean and deepening of the thermocline. The oceanic path occurs along the eastern margin of the basin, where Kelvin waves propagate poleward, depressing the thermocline and resulting in anomalously warm temperatures (Rienecker and Mooers, 1986; Simpson 1983). Increased advection from the south has also been measured directly by current meters (Huyer and Smith, 1985). The atmospheric path is usually slower, resulting in an anomalously strong Aleutian low that may be displaced to the south and/or west and a weaker North Pacific high that may be displaced to the south. The change in atmospheric pressure and storm tracks results in wind-driven currents that transport warmer water onshore. Upwelling in summer does not necessarily weaken but its effects (cold SST and higher pigment concentrations) appear trapped closer to the coast than during other phases of the ENSO cycle (Fiedler, 1984). The boundaries between the regions in the CCS (Figure 1) have been observed to shift during these warm phases causing northward shifts in the distributions of species and their spawning patterns (Pearcy and Schoener, 1987; Wooster and Fluharty, 1985). Onshore and northward shifts of predators create unusually high mortality rates for herring and salmon smolt. Primary production, phytoplankton stocks, and thus food resources for zooplankton are reduced due to reduced nutrient supply. Seabird reproduction in some regions is also impacted by the disappearance of their euphausiid prey (Ainley and Boekelheide 1990). Thus, a number of physical and biological parameters are strongly affected during the warm phase of the ENSO cycle, although the effects may differ among the different regions.
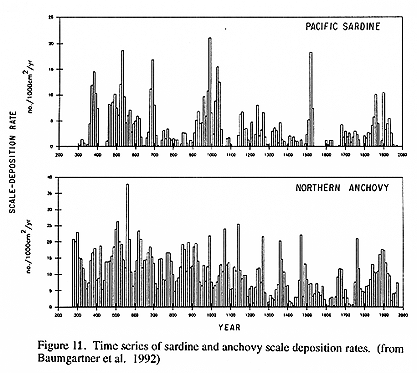
On longer time scales, time series of SST display distinct multidecadal periods of relatively cool and warm conditions, punctuated by the ENSO warm events. SST observations at higher latitudes display a periodicity of about 20 years that may be linked to long period tides (Royer 1993) or atmospheric pressure anomalies (Emery and Hamilton, 1985; Trenberth, 1990). Sediment cores reveal a 60-70 year cycle in the relative dominance of sardine and anchovy over the last 1500 years (Baumgartner et al., 1992) and the direct observational time series (50-100 years) resolve one cycle of this pattern in the physical environment as well as the ecosystem structure-warming around 1925, cooling around 1948 and warming again around 1976 (Hollowed and Wooster, 1992). Shifts from one state to another (warm to cool or vice versa) are often accompanied by concommitant shifts in biological conditions, as abundances, distributions or compositions shift.
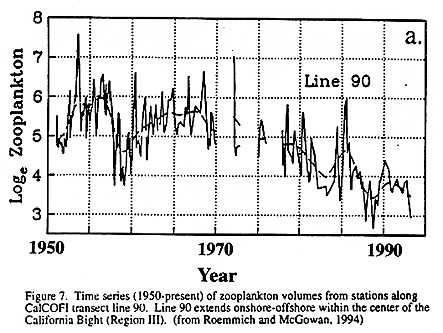
The most recent shift is particularly well documented. Coincident with the change in SST was a notable decrease of zooplankton biomass in Region III (Roemmich and McGowan, 1994) and an increase in the sardine population in Southern California (Barnes et al. 1992). The strength of springtime northerly winds diminished during this period and there is evidence that the large-scale ocean-atmosphere circulation changed after 1976 (Trenberth, 1990; Graham, 1994; Miller et al., 1994b; Trenberth and Hurrel, 1994). From these observed changes, it is reasonable to assume that interdecadal fluctuations in ocean climate, such as those modeled by Miller et al. (1994a, 1994b), create changes in habitat that tend to favor certain species over others on similar time scales. Analysis of the ecosystem response to these interdecadal changes in forcing is especially promising in the CCS, due to the exceptional time series available from the region.
Despite such shortcomings, coupled GCMs may provide broad indications of global warming induced changes in the large-scale Pacific Basin atmospheric and oceanic circulation and internal ocean structure, from which we can hypothesize physical and biological changes in the CCS. For example, coupled GCMs suggest that an atmospheric doubling of CO2 will increase land-sea temperature differences (Trenberth, 1993), intensify equatorward winds in summer and lead to stronger coastal upwelling (Bakun, 1990). Other studies point out that the effects will be more complex and regional than a simple increase in wind strength everywhere (Hsieh and Boer, 1992). Trenberth (1993) discusses implications from the coupled global models and suggests that: (1) the western coast of North America is expected to warm by 1-1.5 deg C by the year 2030; (2) northern regions will warm more rapidly than equatorial regions, decreasing the strength of the mean atmospheric circulation; (3) monsoonal circulation will intensify as the land heats more than the ocean, supporting Bakun's (1990) hypothesis; (4) the hydrologic cycle will strengthen by approximately 10%, implying greater extremes of evaporation and precipitation; and (5) ENSO cycles will continue to exist and to produce greater extremes of flood and drought in the tropics (also found by Meehl, et al., 1993).
On the oceanic side of the coupled GCMs, gradual CO2 doubling causes temperatures in the ocean at mid-latitudes to increase by 2 deg C at the surface, 1 deg C at 500 m and 0.5 deg C at 700 m. Thus, both temperature and stratification increase in the upper water column (Bernal 1993). At higher latitudes, heating is only 1.5 deg C at the surface but it extends deeper. Manabe et al. (1991) show this warming to be slightly less in the NE Pacific than in the zonal mean, with values of 1-2 deg C at the surface.
With respect to forcing by winter storm winds, the model results suggest either no change or a decrease of storm activity at mid-latitudes. Stevensen and Held (1993) show changes in the zonal atmospheric circulation that includes weakening of the subtropical jet on its equatorward side and strengthening of the jet on its poleward side (north of 45 deg N), i.e., a northward shift in the jet. They find the Atlantic storm track to be weakened and imply no change in the Pacific storm track, although they caution against over-interpreting the details of the coarse resolution model results. In fact, many of the authors of the present generation of climate models caution us about the high level of uncertainty that exists in the model results. Keeping this caution in mind, several possible effects of climate change on the circulation and ecosystem dynamics of the CCS can be hypothesized from these model results, in some cases supported by observations.
In regions where the Bakun hypothesis is correct, greater upwelling and increased stratification will create stronger upwelling fronts and jets as well as increased vertical shear within these jets. Since the growth rate of baroclinic instabilities is proportional to vertical shear they should be more unstable, leading to greater mesoscale activity in summer. Finally, through Ekman balance there will be an increase in average offshore transport at the surface, with compensating onshore flow below to feed the increased coastal upwelling. At the same time, mean southward transport in the core of the CCS is predicted to decrease due to: 1) a decrease in the inflow to the CCS from the north as the strength and position of the West Wind Drift is altered by latitudinal-dependent global warming; 2) a decrease in the mean atmospheric circulation and thus ocean forcing; and, 3) little change in storm-driven flow entering along the western open boundary of the EBC region. If thermocline depths and stratification do not change with global warming in part or all of the CCS, the Bakun hypothesis suggests increased nutrient delivery to surface waters which should increase primary production and increase phytoplankton and higher trophic level biomasses.
Peterson et al. (1993), however, argue that global warming will cause the thermocline and nutricline to be depressed along the west coast of the Americas. Through analogy with the warm phase of ENSO, they argue that the net result of increased upwelling-favorable wind stress and a depressed nutricline will be greater upwelling, offshore transport and turbulent mixing, but decreased nutrient concentrations in the mixed layer. Even in the absence of a depressed thermocline in the eastern Pacific, an increase in stratification creates the possibility that much of the onshore flow compensating the offshore surface flow may come from above the nutricline, also reducing nutrient levels in the mixed layer. Reduced nutrient levels in the mixed layer would impact phytoplankton productivity and species composition and perhaps lengthen food chains, resulting in reduced fish biomass.
Storm-driven inflow along the western boundary-Hypothesis: No change or decrease. As discussed above, several factors may affect the strength of storms, changing the strength of the mean northward winds over the western portion of the CCS and the onshore Ekman transport into the CCS. Stephenson and Held (1993) suggest that present models do not indicate a strong change but the weaker overall circulation suggests a decrease in storm winds at mid-latitude.
Outflow to the south-Hypothesis: decrease. By simple continuity, if there is less inflow at the north and no change along the western boundary, the outflow will decrease. This might be slightly altered during summer, if the increased upwelling draws water from beneath the main current into it or if the stronger upwelling produces a stronger jet that can pull more water into the system from the north. During the warm phase of ENSO's, there is a transient northward flow associated with the poleward propagation of the equatorial signal, but there would not necessarily be an analogous northward flow due to gradual heating.
Freshwater input at the coast-Hypothesis: increase in the Pacific Northwest. The increase in the strength of the hydrologic cycle implies an increase in precipitation in the Pacific Northwest, which enters the ocean primarily in the Columbia River and through the Straights of Juan de Fuca. This would slightly increase the stability in the top 1-20 m of the ocean, north of approximately 43 deg N. A change in storm track location and strength could alter this effect (i.e., if more storms are diverted into the Gulf of Alaska).
Timing and persistence of the spring transition-Hypothesis: unspecific change. The occurrence of the spring transition in winds in the Northeast Pacific is part of a rapid large-scale expansion of the North Pacific High, which coincides with a reorientation of the jet stream, similar to a brief (approximately 10 day) blocking event (Strub and James, 1988). This may involve the interaction of the atmospheric circulation and the seasonally developing land-sea temperature difference, with pressures over the land dropping as the continent heats. The weakening of the strength of the atmospheric circulation and the increase in the land-sea temperature difference seems likely to change the strength, timing or even existence of the spring transition, but no guess is offered on the form this change may take.
Mean temperature and stratification-Hypothesis: increase in both. The hypotheses are based on the mean model results, since it is unlikely that the present coarse-resolution models represent the local conditions in the CCS. Given the mean upward slope in the thermocline from west to east, the mean change in temperature and stratification may be more representative of the deeper ocean and thus weaker than that produced by the Stouffer et al. (1989) model. Given the coarse resolution of the model, it is possible that the model misses important differences between the CCS and the mid-ocean.
Transport in alongshore upwelling jets-Hypothesis: increase. Upwelling converts vertical density differences to horizontal differences across a front. If both the ambient vertical stratification and wind-driven coastal upwelling increase, stronger fronts will be created, with stronger alongshore jets. Stronger turbulent mixing may deepen or widen these jets, reducing velocities, but transport will still be greater.
Transport in mesoscale meanders and eddies-Hypothesis: increase. If the previous hypothesis is correct, the stronger horizontal density gradient in the upwelling front implies greater geostrophic shear in the jets. This will make them more unstable, leading to greater meandering of stronger jets and subsequent eddy activity.
Offshore transport-Hypothesis: increase. One of the robust findings of Ekman theory is that the vertically integrated wind-driven transport is in good agreement with the simple Ekman calculation. This has specifically been tested, and confirmed, over the shelf of eastern boundary currents under upwelling conditions (Lentz, 1992). Thus, where Bakun's hypothesis is correct, there will be increased average offshore transport at the surface, with compensating onshore flow below and upwelling along the coast. There will also be increased transport in the offshore portions of the meandering jets, if their strength increases as hypothesized.
Vertical transport (upwelling)-Hypothesis: increase. Upwelling at the coast will increase with increased summer winds, as stated above. Upwelling over the region between the coast and 100-200 km offshore will increase due to increased wind stress curl. Upwelling due to the internal dynamics of eddies and jets will increase with the increase in mesoscale activity. Whether this upwelling draws water from above or below the pycnocline is a crucial question.
Movement of Regional Boundaries-No hypothesis. As stated earlier, one simple prediction for the response of the California Current System is that the boundaries between the physical and biological regions will shift latitudinally. This could occur due to changes in large-scale horizontal advection (north-south) or due to changes in the surface forcing (mostly winds) and the resultant circulation patterns. Movement of these boundaries has been observed during El Ninos, possibly caused by the increase in northward transport, by storm-driven onshore transport of warmer water, or both.
As an example of the kind of changes that could occur if upwelling-favorable winds increase in summer as a result of global warming, we offer this scenario. If the physical boundary between Regions I and II off southern Oregon is caused by the presence of Cape Blanco, the location of this boundary would not be expected to change. The hypothesized increased in wind stress, mesoscale activity and upwelling might, however, make the Southern California Bight look more like Region II, moving the boundary between Regions II and III to the south and changing the nature of the Bight. This could have severe impacts on the pelagic species that presently use the Bight as a nursery. This is offered only as an example, not as a prediction. One of the questions to be answered by the research suggested by this science plan is the extent to which these boundaries have changed in the past during normal climate variations and whether their movement will increase or decrease as a result of global climate change.
Vertical Structure-Hypothesis: Increase in stratification, decrease in mixed-layer nutrients. The vertical structures of temperature, density and nutrients are important parameters for biophysical interactions and offer an example of the complexity of those interactions. Changes in these structures depend upon a mix of factors: 1) the ambient structure of the deep ocean (especially stratification, depth of the thermocline and nutricline); 2) the degree of vertical turbulent mixing (depending on wind mixing power and stratification); and, 3) the strength of upwelling. The predictions of stronger ambient stratification and stronger winds in summer, lead to an increase of upwelling, but changes in the depth of the mixed layer depend on the relative balance of mixing power and stratification. If the increased wind mixing is capable of penetrating the increased stratification and creates a mixed layer as deep as found at present, the overall water column must still be more stable, resulting in a greater density jump at the bottom of the mixed layer. If the wind penetrates farther, the density jump will be even greater. This raises the possibility that much of the onshore flow compensating the offshore surface flow may come from above the thermocline and nutricline, reducing nutrient levels in the mixed layer. Three-dimensional dynamical models of the coastal ocean are needed to determine the effect of specified changes in winds and stratification. Those models must have realistic parameterization of turbulence to determine the effect on nutrient distributions.
Basin-scale circulation patterns may also affect the vertical structure of density and nutrients. Peterson et al. (1993) argue that the basin-wide density structure under global warming scenarios will resemble conditions during the warm phase of ENSO cycles. They argue that: 1) the warm pool in the western Pacific will expand eastward and reduce the fetch over which trade winds travel between the cool eastern tropical Pacific and the warm pool; 2) the reduced fetch (similar to the Atlantic) will result in less inclination of the thermocline from west to east and a deeper thermocline and nutricline in the eastern Pacific; and, 3) the present west to east tilt due to equatorial dynamics extends to at least 30 deg N, implying that deeper thermoclines and nutriclines will be found at temperate latitudes, although less strongly off North America than off South America. If the thermocline and nutricline are depressed by global warming along North America, the net result of increased upwelling-favorable winds will be greater upwelling, offshore transport and turbulent mixing, but decreased nutrient concentrations in the mixed layer, similar to El Nino conditions. They further discuss the implications of these conditions for: 1) recruitment of different species of fish and benthic invertebrates (less favorable for sardine/anchovy); 2) predator/prey interactions (fewer pelagic fish, more euphausiids); and, 3) food web energy dynamics (longer food chains, less fish biomass, and an increased energy supply to the benthos). Although one may disagree with many of their arguments about individual mechanisms or interactions, their discussion serves as an example of the complexity of the interactions and feedbacks involved in determining the complete biophysical response of eastern boundary currents to changing environmental conditions.
{kind=link}
{kind=link}
{kind=link}
{kind=link}