Regional Division
Boundary Shifts
Life History Traits
Genetic Considerations
The four regions of the CCS, defined above from physical attributes, have
many unique biological characteristics. A more detailed and referenced
discussion of these regional differences is found in Section VIII and is
only briefly summarized here.
Region II-Cape Blanco to Point Conception. Primary production is strongly seasonal, and can be elevated in mesoscale jets and eddies. Cape Blanco (to Cape Mendocino) is the approximate southern boundary of Subarctic zooplankton species; in Region II the dominant zooplankton fauna consists primarily of Transition Zone species and no Region II endemics are known. Zooplankton biomass varies seasonally. There are a few major estuaries. Region II is the latitudinal minimum in spawning by epipelagic fishes.
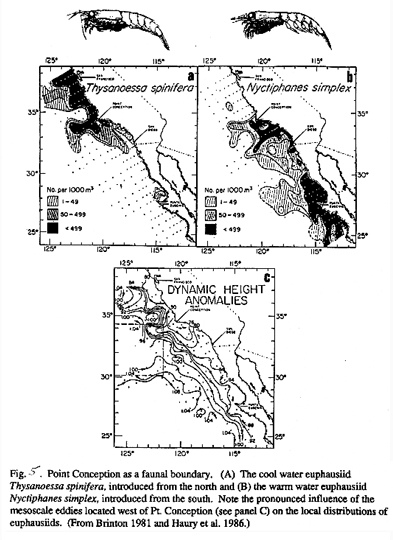
Region III-Point Conception to Punta. Baja. Primary production shows damped seasonal variation, which is often exceeded by interannual variations. The zooplankton is a mixture of Transition Zone species with warm water taxa introduced from the south and the west. An example of Pt. Conception as a distributional boundary for two euphausiid species can be seen in Figure 5. Zooplankton biomass shows relatively small amplitude seasonal variability. Copepods may show both overwintering at depth and continuous reproduction. Region III is the latitudinal maximum in spawning by epipelagic fishes.
Region IV-Punta Baja to Cabo San Lucas, Southern Baja California. There is modest seasonality in primary production, much of which may be localized in major upwelling centers. The zooplankton assemblage is a mixture of some Transition Zone and other cool water species with Equatorial and Eastern Tropical Pacific fauna. Zooplankton biomass shows damped seasonality. Fish spawning tends to be geographically localized.
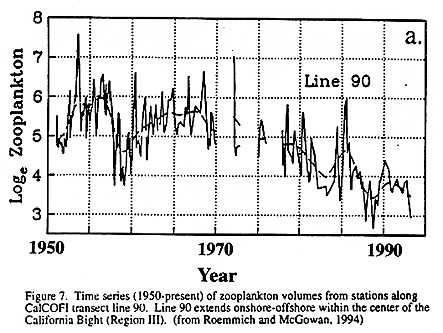
Decadal Scale Shifts. Major biological shifts in the CCS have been documented for the recent warm period (beginning in 1977; see Fig. 9), including a five-fold drop in zooplankton abundance within the Southern California Bight (Roemmich and McGowan 1994; Fig. 7) and recovery of depleted Pacific sardine stocks (Barnes et al. 1992). We do not know how these are related to spatial boundaries in the CCS. However, during this same period, zooplankton biomass in the subarctic Pacific has doubled (Brodeur and Ware 1992) as has that of squid (Brodeur, in prep), suggesting major shifts in productivity of the subarctic gyre (increased) and California Current (decreased) during the present warm regime.
Longer term, interdecadal scale variations in zooplankton abundance in parts of Region III of the CCS have also been observed. Roemmich and McGowan (1994; Fig. 7) document a long term decline in zooplankton biomass within the Southern California Bight that began approximately at the time of the mid-70's warming event in the CCS.
Sedentary adults of marine benthic invertebrates clearly cannot undertake the spawning migrations of motile fishes. Although spawning of marine benthic invertebrates may not be localized to one region of the CCS, recruitment success to the nearshore marine benthos can be. Roughgarden et al. (1988), for example, suggest that the recruitment success of intertidal barnacles along the central California coast is strongly influenced by cross-shelf transport of larvae, which may be controlled by onshore-offshore displacement of alongshore fronts. Recruitment is favored when the fronts are nearshore and act as a barrier to offshore dispersal. In Region I, however, where cross-shore transport is less intense (and onshore flow associated with downwelling occurs for part of the year), the supply of larvae is thought to be capable of saturating benthic substrates. Where the supply of larvae is saturating, adult-adult interactions or predation on the bottom may be the most important determinants of population growth and benthic community composition (Paine, 1984). Similarly, topographically controlled nearshore circulation can regulate recruitment success both in space and time. Recruitment of purple sea urchin (Strongylocentrotus purpuratus) larvae to the benthos is influenced by latitudinal variations in the presence of topographic features such as capes and headlands. Such features tend to be sites of strong offshore transport and thus reduced nearshore larval supply, resulting in infrequent events of successful recruitment (Ebert and Russell, 1988). Wing et al. (in press) have shown how upwelling relaxation can lead to northward, onshore transport and subsequent recruitment of crab species in Region II. Concentration of larvae in upwelling shadows in the lee of promontories, with subsequent northward transport may provide a retention mechanism in the face of stron southward currents during upwelling.
Satellite images reveal that zones of enhanced phytoplankton biomass extend further offshore in Region II than elsewhere in the CCS, in a rich variety of mesoscale features including eddies, jets, and coastal filaments (Strub et al., 1991). While experimental evidence suggests that copepod lipid reserves and egg production may increase in response to the enhanced food supply in coastal filaments (Smith and Lane, 1991), the ultimate fate of this elevated copepod secondary production is not known.
The life history traits of sibling species of marine copepods differ in the north and south of the CCS. Calanus pacificus, for example, shows seasonal diapause at depth in Region I (in Puget Sound, Osgood and Frost, 1994), but in Region III only one fraction of the C. pacificus population undergoes diapause (Longhurst, 1967; Alldredge et al., 1984) while another fraction shows continuous year-round reproduction in near-surface waters (Mullin and Brooks, 1967). Similar differences in diapause response are known for a population within a region: the Calanus marshallae population in Dabob Bay, a fjord at the northern end of Region I, produces a single generation which begins in February and goes into diapause in May. The population which resides in shelf waters off Oregon and Washington (also Region I) passes through multiple generations, beginning in January/February and extending through September.
Comparative studies of conspecifics in different parts of their range are likely to lead to improved understanding of both the proximate cues and ultimate advantages of different life history traits. These studies will provide the basis for the development of predictive models of population growth under different scenarios of a changing ocean climate.
Isolation of subpopulations, particularly in regions of steep environmental gradients, can lead to genetic differentiation of populations through local adaptation or genetic drift. There are clearly large environmental gradients among regions of the CCS, however, subpopulations of most marine species are unlikely to be isolated from one another due to the large scale dispersal of planktonic stages. The ensuing gene flow associated with this dispersal should homogenize the genetic composition of populations over large regions (Burton 1983, Gyllensten 1985, Hedgecock 1994a). Comparisons of life histories and vital rates for many widespread species have found significant differences among regions of the CCS. U.S. GLOBEC studies must determine if genetic variation contributes to this geographical variation in key traits. Genetic studies will need to resolve whether species thought to range widely in the CCS are indeed single species or instead sibling species, whose morphological similarity masks underlying genetic differences. Similarly, population genetic studies will need to characterize the amount of spatial genetic variation within species. Such studies could reveal exceptions to the expectation of spatial homogeneity (e.g., some evidence exists for differentiation of northern and southern populations of Calanus; Bucklin and Kann, 1991), or they could reveal previously unrecognized barriers to dispersal (e.g., Avise et al. 1987, Burton 1994).
Temporal genetic change via recruitment appears to be as large or larger than geographic variation on broad spatial scales (Johnson and Black, 1984; Hedgecock, 1994a, 1994b). Molecular methods could be applied to preserved zooplankton (such as in the CalCOFI collections) to establish time-series estimates of genetic structure and variation for targeted taxa. Moreover, population genetic studies are also important for estimating effective population size-i.e., an estimate of the number of individuals that successfully reproduce-which has recently been shown to be surprisingly small, often many orders of magnitude less than adult abundance (Hedgecock, 1994b). If so, this establishes a link between oceanographic processes and population genetic structure which might be exploited to help explain how local adaptations and speciation can occur in seemingly large, well-mixed marine populations.
Understanding how widely distributed species respond evolutionarily to climatic variation over a broad geographic range may provide important clues to predict how distributions may be altered by climate change. It is equally important, however, to understand how more narrowly distributed species may respond to climate change, because they may respond first or be most seriously affected by future climate change.
{kind=link}
{kind=link}
{kind=link}