Phytoplankton Provinces
Zooplankton Provinces
Invertebrate Benthos Provinces
Marine Fish Provinces
Bird Provinces
El Nino Effects on Zoogeographic Boundaries
Many species exhibit distinct distributional boundaries that coincide
with changes in the regional oceanographic conditions described
previously. These boundaries are indicators of responses of marine
organisms to key physical factors and may provide clues to the principal
processes underlying production in the CCS. A brief description of
these provinces follows.
Local oceanographic conditions appear to influence the distribution of phytoplankton. Tont (1976, 1981) showed diatom abundances to be associated with coastal upwelling, with several blooms of 5-6 weeks duration during summer. Hood et al. (1990, 1991) and Abbott et al. (1990) showed Chaetoceros and Skeletonema costatum to be the dominant diatoms in several chlorophyll maxima in the CTZ region. Their distribution, however, was patchy, with Rhizosolenia alata, Thalassiosira spp. and Nitzschia spp. dominating in a tongue of cold water extending seaward from Cape Mendocino.
Shifts in phytoplankton species dominance occur over alongshore distances of less than 100 km and time periods of days. Abbott et al. (1990) found that species composition along a drifter track changed over time scales of 2-3 days. This change was ascribed to in situ growth of large centric diatoms (probably Actinocyclus or Thalassiosira) in newly upwelled water. Concentrations of nitrate+nitrite dropped from 10 micromolar to zero during this time, while chlorophyll decreased and cell volume increased.
The Transition Zone fauna (e.g., copepods Eucalanus californicus, Pleuromamma borealis; euphausiids Nematoscelis difficilis, Thysanoessa gregaria; chaetognath Sagitta scrippsae) can extend from British Columbia to the tip of Baja California, thus spanning Regions I-IV. Notable among the euphausiids is Euphausia pacifica which inhabits both the Subarctic and the Transition Zone, with centers of abundance in both the Gulf of Alaska and off central and southern California (Brinton 1962).
The population dynamics and phenology of some zooplankton species which span Regions I-III have been examined and found to differ between regions, perhaps due to adaptations to local environmental conditions. For example, although seasonal variations in wind stress, upwelling indices, and particulate organic matter are detectable in Region III (Brinton 1976, Mullin 1986) seasonal variations in primary production rates are relatively damped and sometimes greatly exceeded by interannual variations (Mullin 1986). The seasonal cycle of zooplankton biomass, measured as displacement volume, has a lower amplitude in Region III than in regions of the CCS further to the north (Roesler and Chelton 1987). The life history characteristics and reproductive responses of zooplankton in Region III are correspondingly less seasonal. The copepod Calanus pacificus has been found to reproduce year-round in Region III (Mullin 1991), while egg production stops in winter in Puget Sound within Region I (Runge 1985). Similarly there is some year-round recruitment of older copepodid stages of C. pacificus in Region III (Mullin and Brooks 1967), while population growth is quite seasonal in higher latitudes with overwintering accumulations of copepodid stages IV and V.
Within individual species of zooplankton, the population genetic structure may correspond to different water types or portions of coastal and upwelling hydrodynamic features (Bucklin 1991). C. pacificus sensu latu is widely distributed and abundant in Regions I-III of the CCS, but appears to have subspecies that are associated with either the Subarctic Pacific in Region I (C. pacificus oceanicus Brodsky) or with Regions II and III (C. pacificus californicus Brodsky). Recent preliminary mitochondrial DNA studies (Bucklin and Kann 1991) have found genetic differentiation between northern and southern populations of C. pacificus, confirming that regional differentiation of closely related species has occurred within the CCS.
There is no defined faunistic boundary for zooplankton between Regions III and IV; some Transition Zone and other cool water zooplankton species occur in the upwelling zone off Punta Eugenia at 27 deg N. The Equatorial and Eastern Tropical Pacific zooplankton faunas can extend northward from lower latitudes along the coast of Baja California into Region IV.
Recurrent patterns of cross-shelf zonation are known for copepods off Washington and central Oregon (Peterson et al. 1979; Peterson, 1980): several species are almost completely restricted to the nearshore zone, out to water depths not exceeding 50 m (Acartia hudsonica, Centropages abdominalis and the cladocerans Evadne nordmanni and Podon leukartii), other species are found predominantly in mid-shelf and slope waters (A. longiremis and Calanus marshallae) and one ranges across the entire domain (Pseudocalanus mimus). Behavioral mechanisms which lead to cross-shelf zonation and retention in the Oregon upwelling zone are discussed in Peterson et al. (1979) for five copepod species, and Peterson (1980). Attempts to model retention mechanisms proposed by Peterson et al. (1979) using a physical transport model were only partially successful (Wroblewski 1980, Wroblewski 1982) and should be revisited.
Cross-shelf zonation does not seem to be observed off central or southern California. Coastal/shelf species such as C. pacificus, Pseudocalanus mimus, Paracalanus parvus, Acartia tonsa and A. longiremis are common hundreds of kilometers seaward of the shelf. Total zooplankton biomass is high out to several hundred kilometers from shore as well (Bernal and McGowan 1981). The apparent lack of zonation off California as compared to Oregon may reflect the fact that upwelling off Oregon is restricted to the shelf whereas off California the area affected by upwelling is far more broad and includes Ekman upwelling nearshore, Ekman pumping offshore and offshore transport of upwelled water via filaments, jets and eddies.
The CCS contains fish common to several zoogeographic provinces. Allen and Smith (1988) defined zoogeographic provinces for the marine fish common to the North Pacific. Allen and Smith (1988) also defined life zones to describe the principal location in the water column that adult fish occupy (Table 6). Based on these definitions it appears that Region II may represent a transition zone between subtropical and subarctic fish species (Tables 2-5). Further south, Parrish et al. (1981) noted that Magdelena Bay (25 deg N) is the average location of a front that forms the boundary between tropical and subtropical fauna.
In general, rockfish, flatfish, and groundfish all exhibit regional distributions that appeared to have either northern or southern boundaries in Region II (Tables 2-5), reflecting two major provinces--a northern subarctic and a southern subtropical. Out of all of the species considered, only ten species inhabit all four regions. These ten species all exhibited pelagic or mesopelagic behaviors (Table 4). Of these, the northern anchovy and Pacific sardine include separate spawning stocks in the north, central and southern portions of the CCS (Parrish et al 1989, Methot 1989).
The zoogeographic boundary of many marine fish stocks may be located in Region II because ocean conditions there may not be favorable to spawning success. As noted by Parrish et al. (1981), few fish species spawn in the surface waters of Region II. Those species that do reproduce there tend to employ life history traits (e.g., live-bearing, spawning in estuaries, anadramous spawning) that minimize the risk of offshore transport in open ocean waters. Hence Region III, and to some extent Region I, are major centers of fish reproduction and of potential fish-zooplankton interactions in the CCS. Region III is the preferred spawning site for over 90% of the epipelagic fish biomass (hake, sardine, anchovy) in the southern part of the CCS. Pacific hake (Merluccius productus) are particularly notable because they undertake a migration of approximately 2,000 km from Vancouver Island to spawn in the offshore waters of Region III (Bailey et al. 1982). At the top of the trophic pyramid, the white shark Carcharodon carcharias displays a similar seasonal and migratory cycle: breeding and pupping in Region III, then moving north to Regions I and II.
Circumstantial evidence suggests that the distribution of some marine birds may be determined by key prey items in the diet. For example, Thysanoessa spinifera is a critical prey item in the diet of Cassin's Auklet. Vermeer (1981) suggests that the geographic distribution of this euphausiid in the eastern North Pacific determines that of the auklet-both T. spinifera and Cassin's Auklet extend from southeast Alaska to northern Baja California, Mexico.
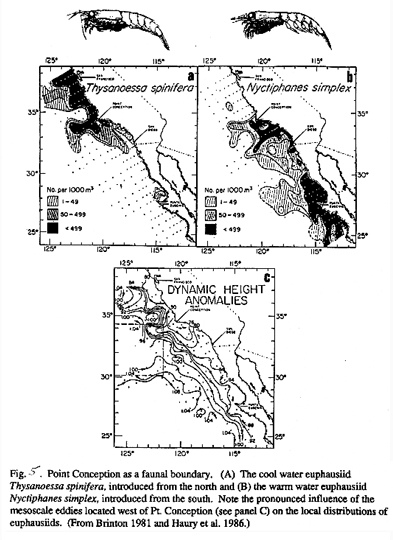
Geographic domains typically inhabited by faunal assemblages may shift northward during extreme ENSO events (Fulton and LeBrasseur 1985, cf. Ware and McFarlane 1989). During the 1983 El Nino, inshore waters off Oregon were dominated by southern copepods, Paracalanus parvus and Acartia tonsa, and there were many occurences of all life stages of the euphausiid Nyctiphanes simplex-previously recorded only as far north as central California (Fig. 5; Miller et al. 1985). Brinton (1967) noted that the southern euphausiid (Euphausia eximia) extended its range north in coastal waters during warm periods. The zoogeographic boundary of some fish species is altered during El Nino events (Pearcy and Schoener 1987, Arntz et al. 1991). Bailey and Incze (1985) noted that El Nino effects appear to favor stocks at the northern end of their range with opposite effects on stocks at the southern end of their range. They also predict that the impact of ocean warming will have the greatest influence on sedentary species and species that exhibit homing or slow migrations. Increased mortality rates for key prey species may be observed when pelagic fish predators shift their distributions. If a prolonged change in ocean conditions occurred, local species assemblages may change due to competition for space and resources, resulting in shifts in species dominance. Moreover, range extensions can have significant impacts on trophic relations, as evidenced by Pearcy et al.'s (1985) observation that Nyctiphanes simplex was the most common euphausiid prey of juvenile coho salmon off Oregon and Washington in 1983. Understanding the potential response of marine stocks to climate change would help to predict future community structures in the CCS.
{kind=link}